
 Kristen A. Lindquist, Department of Psychology, University of North Carolina, Chapel Hill
Kristen A. Lindquist, Department of Psychology, University of North Carolina, Chapel Hill
It goes without saying that the brain produces emotions—in this day and age, you’d have to be a pretty staunch dualist to argue otherwise. The big question that remains concerns how the brain creates emotions. Historically, it was assumed that each emotion had a discrete biological core responsible for its creation, such as a certain brain area (e.g., the amygdala) or a network of areas in the evolutionarily “old” portion of the brain (e.g., a network in the brainstem and other subcortical regions). This idea is known as faculty psychology (Lindquist & Barrett, 2012) —it is the commonsense notion that specific experiences (anger, disgust, fear, thinking, remembering, acting) each correspond to a specific, evolved, and anatomically defined neural mechanism.
In contrast to this faculty psychology approach, recent research suggests that emotions are instead constructed out of more fundamental psychological processes that are not themselves specific to emotion (for reviews see Barrett, 2006; Barrett, 2009; Cunningham et al., 2013; Lindquist, 2013; Lindquist & Gendron, 2013; Lindquist, Wager, Kober, Bliss-Moreau & Barrett, 2012). In this view, instances of emotion (e.g., an experience of anger at a friend) emerge from basic, interacting psychological ‘ingredients’ that each perform a domain-general psychological function that contributes to a variety of emotions, cognitions, perceptions, and actions.
In the constructionist approach my colleagues and I take to understanding the nature of emotion (Barrett & Satpute, 2013; Lindquist, 2013; Lindquist & Barrett, 2012; Oosterwijk et al., 2012; Wilson-Mendenhall et al., 2011), we hypothesize that these basic ingredients involve representations of the body (called “interoceptive sensations” or “core affect”), representations of prior experiences (called “concept knowledge”), representations of the external world (“exteroceptive sensations”) and attention (“executive control”). According to our approach, the brain is a predictive system (see Friston & Kiebel, 2009), always using prior experiences to disambiguate and make meaning of sensations from the body and world. A person experiences an emotion (e.g., anger at a friend) when he makes a “situated conceptualization” of his core affective state in a given context using concept knowledge about emotion (e.g., knowledge about the concept of anger, prior experiences of anger in a similar context)”.
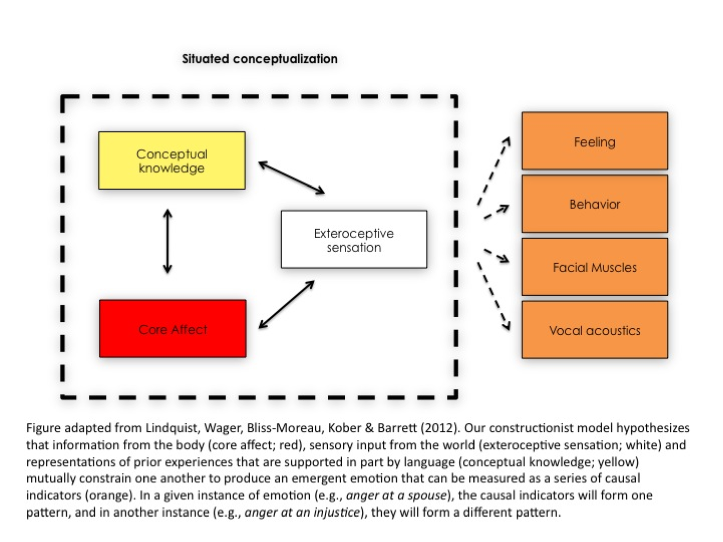
Growing neuroscientific evidence from human lesion studies, neuroimaging, and studies of non-human animals supports such a constructionist view of the mind; emotions appear to involve the interaction of neural networks that serve domain-general functions that are not specific to emotions. Contrary to a faculty psychology view, the empirical evidence suggests that certain brain regions are not specific to certain emotions. For instance, although once heralded as the brain basis of fear in mammals, research shows that the human amygdala has neither consistent nor specific increases in activation during instances of fear. Individuals with amygdala lesions can still perceive fear on others’ faces when they are specifically directed to look at the diagnostic features of fearful faces (i.e., the eyes; Adolphs et al., 2005). Individuals with amygdala lesions can even experience intense fear when deprived of oxygen (Feinstein et al., 2013). Nor is the human amygdala specific to fear. During neuroimaging experiments of healthy individuals, it shows increased activity during the experience and perception of many different emotions (Lindquist et al., 2012; Vytal & Hamann, 2010).
Another important source of evidence in favor of a constructionist view of the mind is the finding that human emotions involve networks that are not themselves specific to emotion. Brain regions demonstrating increased activity during emotion experiences and perceptions appear to play core affective, conceptual, sensory, and executive control roles across other psychological domains such as moral judgments, empathy, autobiographical memory, and even visual perception (Lindquist & Barrett, 2012).
These networks interact with one another when a person experiences an emotion, consistent with the idea that emotions emerge from the combination of domain-general networks, rather than from a single emotion-specific network. For instance, a brain network supporting core affect shows increased interaction with a network supporting conceptualization as the intensity of sadness increases over the course of a movie (Raz et al., 2012). Complex patterns of activity within these interacting domain-general networks are also associated with experiences of different emotion categories (e.g., anger, disgust, fear, lust, etc.) (Kassam et al., 2013).
Although it is tempting to assume that evidence for such patterns across brain networks involved in core affect, conceptualization, attention and sensory perception is evidence for the evolutionarily given circuit for an emotion category, it is problematic to interpret neuroscientific data in this manner. First, the pattern observed for a certain emotion category (e.g., anger) differs based on the context in which it is experienced (e.g., a physical v. a social situation; Wilson-Mendenhall et al., 2011), suggesting there is no single network for that emotion category. Second, scientists also observe patterns of brain activity that correspond to experiences of nominal kind categories that are human constructions learned through experience such as cars, bottles, and athletes (Huth et al., 2012). But we would never assume that a neural pattern associated with perceiving a car gives evidence for the evolutionarily-endowed “car network”. Consequently, it’s not clear why similar data would provide evidence for an evolutionarily-endowed “fear network.” The evidence is thus more consistent with the idea that emotions emerge from the combination of more basic neural parts that perform domain-general functions.
A further piece of evidence in favor of a constructionist perspective is that the domain-general networks that support basic psychological functions such as core affect, conceptualization, exteroceptive sensation and executive attention appear to be “intrinsic” networks that are constrained by anatomical connections between brain areas (e.g., Yeo et al., 2011). Many of these networks exist in non-human animals (Rilling et al., 2007; Vincent et al., 2007) and develop in humans across early life (Gao et al., 2011). We thus suggest that they comprise a set of basic functional building blocks of the mind.
In contrast to these domain-general intrinsic networks, neural networks that support adaptive mammalian behaviors (e.g., freezing, attack, maternal behavior; Panksepp, 2004) are sometimes cited as emotion-specific networks. Many of these largely subcortical networks are indeed preserved across species and are certainly relevant to human emotions. Yet neither human nor non-human emotions seem reducible to the specific behaviors supported by such networks—a network for freezing, for instance, cannot logically be considered the network for a complex category such as fear since both humans and non-human animals engage in many behaviors beyond freezing in the face of a threat (Barrett et al., 2007; LeDoux, 2012).
If we reduce the category of fear to the network for freezing, we cannot say that a human or even a rat is fearful when it attacks a threatening intruder, flees, or engages in other defensive behaviors. Instead, networks supporting these adaptive mammalian behaviors can be considered additional ‘ingredients’ of the mind that contribute to some, but not all instances of emotion. For example, a network for freezing might contribute to an instance of fear when a person sees a snake in the garden, but not when she strikes a mugger in a dark alley.
To sum up, growing evidence demonstrates that a constructionist approach is a useful avenue to understanding emotion-brain correspondence. What remains to be seen is whether the ingredients hypothesized at present (e.g., Lindquist & Barrett, 2012) are the best candidates for the basic ‘ingredients’ of the mind more generally. As network-based neuroscientific approaches progress, we might find that alternative, more specific formulations of intrinsic networks are in fact the best candidates for the brain’s functional building blocks. This process of discovery will rely not just on advanced technology, but also on precise and well-validated psychological models of emotion that can constrain the interpretation of neuroscientific results.
References
Adolphs, R., Gosselin, F., Buchanan, T. W., Tranel, D., Schyns, P., & Damasio, A. R. (2005). A mechanism for impaired fear recognition after amygdala damage. Nature, 433, 68-72.
Barrett, L. F. (2006). Are emotions natural kinds? Perspectives on Psychological Science, 1, 28-58.
Barrett, L. F. (2009). The future of psychology: Connecting mind to brain. Perspectives on Psychological Science, 4, 326-339.
Barrett, L. F., Lindquist, K. A., Bliss-Moreau, E., Duncan, S., Gendron, M., Mize, J., et al. (2007). Of mice and men: Natural kinds of emotions in the mammalian brain? A response to panksepp and izard. Perspectives on psychological science: a journal of the Association for Psychological Science, 2, 297-297.
Barrett, L. F., & Satpute, A. B. (2013). Large-scale brain networks in affective and social neuroscience: Towards an integrative functional architecture of the brain. Current Opinion in Neurobiology.
Cunningham, W. A., Dunfield, K. A., & Stillman, P. E. (2013). Emotional states from affective dynamics. Emotion Review, 5, 344-355.
Feinstein, J. S., Buzza, C., Hurlemann, R., Follmer, R. L., Dahdaleh, N. S., Coryell, W. H., et al. (2013). Fear and panic in humans with bilateral amygdala damage. Nature Neuroscience, 16, 270-272.
Friston, K., & Kiebel, S. (2009). Predictive coding under the free-energy principle. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 364, 1211-1221.
Gao, W., Gilmore, J. H., Giovanello, K. S., Smith, J. K., Shen, D., Zhu, H., et al. (2011). Temporal and spatial evolution of brain network topology during the first two years of life. PLOS One, 6, e25278.
Huth, A. G., Nishimoto, S., Vu, A. T., & Gallant, J. L. (2012). A continuous semantic space describes the representation of thousands of object and action categories across the human brain. Neuron, 76, 1210-1224.
Kassam, K. S., Markey, A. R., Cherkassky, V. L., Loewenstein, G., & Just, M. A. (2013). Identifying emotions on the basis of neural activation. PLOS One, 8, e66032.
LeDoux, J. (2012). Rethinking the emotional brain. Neuron, 73, 653-676.
Lindquist, K. A. (2013). Emotions emerge from more basic psychological ingredients: A modern psychological constructionist model. Emotion Review, 5, 356-368.
Lindquist, K. A., & Barrett, L. F. (2012). A functional architecture of the human brain: Insights from emotion. Trends in Cognitive Sciences, 16, 533-554.
Lindquist, K. A., & Gendron, M. (2013). What’s in a word: Language constructs emotion perception. Emotion Review, 5, 66-71.
Lindquist, K. A., Wager, T. D., Kober, H., Bliss-Moreau, E., & Barrett, L. F. (2012). The brain basis of emotion: A meta-analytic review. Behavioral and Brain Sciences, 35, 121-143.
Oosterwijk, S., Lindquist, K. A., Anderson, E. C., Dautoff, R., Moriguchi, Y., & Barrett, L. F. (2012). States of mind: Emotions, body feelings, and thoughts share distributed neural networks. NeuroImage, 2110-2128.
Panksepp, J. (2004). Affective neuroscience: The foundations of human and animal emotions: Oxford University Press, USA.
Raz, G., Winetraub, Y., Jacob, Y., Kinreich, S., Maron-Katz, A., Shaham, G., et al. (2012). Portraying emotions at their unfolding: A multilayered approach for probing dynamics of neural networks. Neuroimage, 60, 1448-1461.
Rilling, J. K., Barks, S. K., Parr, L. A., Preuss, T. M., Faber, T. L., Pagnoni, G., et al. (2007). A comparison of resting-state brain activity in humans and chimpanzees. Proceedings of the National Academy of Sciences of the United States of America, 104, 17146-17151.
Vincent, J. L., Patel, G. H., Fox, M. D., Snyder, A. Z., Baker, J. T., Van Essen, D. C., et al. (2007). Intrinsic functional architecture in the anaesthetized monkey brain. Nature, 447, 83-86.
Vytal, K., & Hamann, S. (2010). Neuroimaging support for discrete neural correlates of basic emotions: A voxel-based meta-analysis. Journal of Cognitive Neuroscience, 22, 2864-2885.
Wilson-Mendenhall, C. D., Barrett, L. F., Simmons, W. K., & Barsalou, L. W. (2011). Grounding emotion in situated conceptualization. Neuropsychologia, 49, 1105-1127.
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D., Hollinshead, M., et al. (2011). The organization of the human cerebral cortex estimated by functional connectivity. Journal of Neurophysiology, 106, 1125–1165.



Can you put a date on this interview for citation purposes?