

November 2014 – Unveiling brain networks that mediate music-induced emotions, or musical emotions, is challenging but of great interest for several reasons. First, musical emotions represent a remarkable and intriguing experience that may yield precious insights into the workings of the human mind. Second, musical emotions lack the sort of obvious adaptive function and cultural invariance characteristic of basic emotions, preventing a direct appeal to evolutionary explanations for their origin. Third, and relatedly, because musical emotions are very different from affective phenomena traditionally investigated in neuroscience, such as fear, joy, disgust, and so forth, they offer useful avenues to study emotions across a wider range and thus compare their neural underpinnings across more diverse samples of affective subjective experiences.
Fourth, studying the brain mechanisms of musical emotions can help us understand the neurocognitive processes at play in the perception and production of music itself. Still another reason is that building bridges between brain science and musical art might eventually give composers or performers novel tools to exploit in artistic creation (conversely, neuroscientists can gain new insights from the work of musicologists and musicians). Of course, these are just a few of the reasons to be interested in musical emotions, and many others certainly apply.
Arguably, neuroscience research has yet to fully exploit the richness and originality offered by emotions in the music domain. A large majority of studies has investigated brain activation patterns associated with dichotomous categories such as pleasant/positive vs. unpleasant/negative emotions. This “standard” approach parallels classic neuroscience work on “basic” emotions and primary reinforcers, such as reward and fear (Armony & Vuilleumier, 2013). However, it does not cover the whole range of emotional experiences in the music domain, which prominently include awe, wonder, and sublimity – i.e., feelings that are often considered as belonging to a special category of “aesthetic emotions” (Zentner, Grandjean, & Scherer, 2008). Conversely, some of the emotions commonly experienced in non-musical contexts – e.g., fear, anger, or even plain happiness – are not frequently experienced when listening to music. Therefore, classic dichotomous distinctions between pleasant/unpleasant or between appetitive/aversive affective dimensions may prove less informative about the neural substrates of musical emotions than about other emotion types investigated in neuroscience.
Another limitation is that many studies using brain imaging techniques such as fMRI or PET have focused on activation patterns associated with emotions during relatively long period of time, typically averaging the brain response over music segments of 20-60 seconds. Although this slow time scale might fit aesthetic emotions better than more basic emotions (such as fear or disgust), it is likely to miss important dynamic features linked to the rich temporal structure of music (such as mixture or rapid succession of different emotions, or expectations built by progression and repetition in music scores). Other neuroimaging techniques with higher temporal resolution, such as electro-encephalography (EEG) or magneto-encephalography (MEG), have been used to examine responses to transient and elementary musical features (e.g. linked to rhythmic, harmonic, or syntactic structure), but surprisingly less often to investigate musical emotions, perhaps reflecting the difficulty to precisely define the onset and duration of these emotions.
All these caveats aside, neuroscience research has already provided abundant data illuminating several key aspects of music-induced emotions (for review, see Koelsch, 2014). A number of pioneering and elegant studies using PET and subsequently fMRI have consistently shown that pleasurable music activates brain regions usually responding to other pleasures and rewards such as the ventral striatum, but also orbitofrontal cortex, anterior insula, together with frequent activations in parietal and somatosensory areas (Blood & Zatorre, 2001; Menon & Levitin, 2005). Similar activations in striatum are found for music evoking joy and happiness (Mitterschiffthaler, Fu, Dalton, Andrew, & Williams, 2007); sad music, on the other hand, has been reported to activate hippocampus, amygdala, and neighboring medial temporal lobe areas, which are implicated in negative affective states and anxiety. Comparing unpleasant dissonant to pleasant consonant music produces similar effects (Blood, Zatorre, Bermudez, & Evans, 1999; Koelsch, Fritz, DY, Muller, & Friederici, 2006).
Furthermore, elegant work combing pharmacological PET with fMRI has demonstrated that transient peaks of pleasure experienced during music correlate with release of dopamine in ventral regions of the striatum (nucleus accumbens), distinct from more dorsal sites of dopamine release associated with anticipation of the corresponding musical moments (Salimpoor, Benovoy, Larcher, Dagher, & Zatorre, 2011). Feelings of “chills” triggered by one’s preferred music also correlate with increased activity in both striatum and insula, accompanied by transient reductions in activity in amygdala, anterior hippocampus, and ventromedial prefrontal cortex (Blood & Zatorre, 2001).
Furthermore, although fear may not be truly evoked in listeners, it can be expressed by music (as in horror movies) and often produce increased activity in the amygdala (Aube, Angulo-Perkins, Peretz, Concha, & Armony, 2014). Interestingly, these amygdala responses overlap with those to other stimuli (such as faces or voices) evoking similar emotions (Aube, et al., 2014), and damage to the amygdala may impair recognition of fear as expressed both in music and faces (Gosselin, Peretz, Hasboun, Baulac, & Samson, 2011). Altogether, these findings have been taken as evidence that music can evoke “true” emotions (rather than more “abstract” or “intellectual” states of mind) – although such conclusion is partly fraught by a problem of “reverse inference” (Poldrack, 2006). Indeed, activity in the limbic system (including amygdala) does not necessarily equate with emotional experience and amygdala activity is often elicited without any reportable feeling (e.g. in response to subliminal emotional stimuli or visible non-emotional stimuli such as eyes).
In contrast, research in psychology has often emphasized a different kind of aesthetic emotions elicited by music, beyond basic emotions such as joy, sadness, and fear, or dichotomous distinctions between pleasant and unpleasant emotions. Based on ratings made by large groups of participants exposed to various genres of music, with more than 500 emotion terms, Marcel Zentner and Klaus Scherer (2008) proposed a set of “music-specific” emotions corresponding to those most commonly reported during music listening across genres. This model distinguished 9 main categories of emotions commonly experienced while listening to music: joy, sadness, tension (fear or worry), wonder, nostalgia, tenderness, power, peacefulness, and transcendence.
Together with Wiebke Johanna Trost, we recently set out to investigate the neural underpinnings of these complex, possibly “music-specific” emotions (Trost, Ethofer, Zentner, & Vuilleumier, 2012). Although commonly reported by music listeners, aesthetic emotions (such as wonder or nostalgia, etc.) have not been systematically studied in brain research. Moreover, rather than applying dichotomous comparisons by contrasting pairs of pre-selected pieces (e.g. pleasant vs unpleasant, joyful vs sad), we designed a parametric approach allowing participants to report different emotions with different intensities for the same music piece. By analyzing subjective ratings for several excerpts from the classic repertoire, we found that the 9 emotions described by Zentner and colleagues (2008)
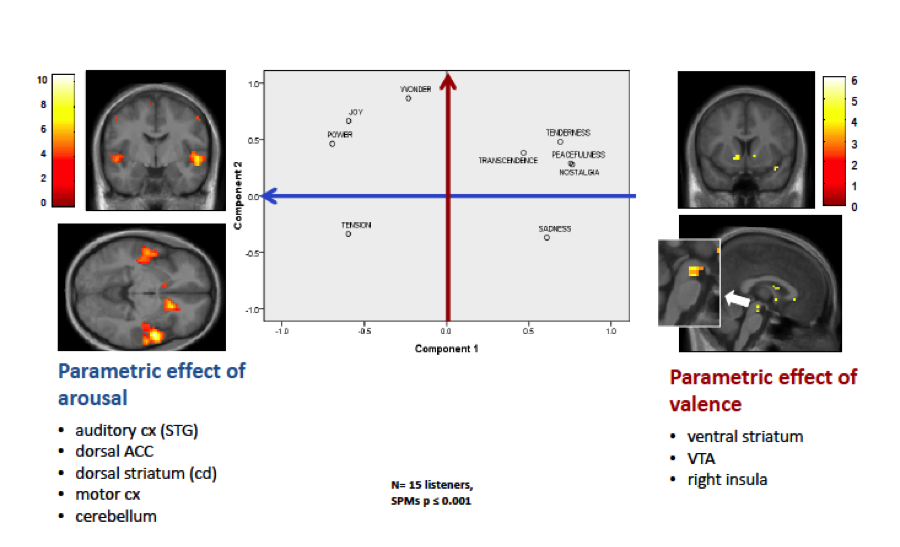
arousal axis correlate with activation in auditory cortex, but also anterior cingulate, premotor cortex, and more dorsal regions in striatum (caudate), all regions associated with vigilance and action
were distributed in a bidimensional space corresponding to the typical organization of valence (from negative to positive) and arousal (from calmness to excitement) (Russell, 2003; see Figure 1). This “circumplex” has often been described as defining the major ingredients of a variety of emotions, including basic categories, across various stimulus modalities (faces, voices, etc.) (Russell, 2003).
Brain activity measured by fMRI in our participants showed that higher ratings along the positive valence axis correlated with selective activations in dopaminergic pathways (ventral striatum and tegmental area) and insula . Conversely, higher ratings of arousal correlated with activation in auditory cortex, but also anterior cingulate, premotor cortex, and more dorsal regions in striatum (caudate), all regions associated with vigilance and action (Figure 1). However, emotions within the same class of valence or arousal were distinguished by variations in the degree of activation of these two basic networks, in combination with differential recruitment of other brain regions.
For instance, relative to joy (which was associated with verbal labels referring to dancing and elation, see Zentner et al. 2008), wonder was associated with activity in striatum, but weaker increases in motor related areas and greater increases in memory related areas including hippocampus. Conversely, tension (encompassing feelings of worry and uneasiness) was accompanied by activation in cortical and subcortical motor pathways, similar to joy, but not in ventral striatum. On the other hand, hippocampal regions and medial prefrontal areas usually involved in self-reflective processes and introspection were activated during positive emotions with low arousal such as nostalgia and tenderness, but not during sadness, where selective increases instead were seen in medial parietal areas and subgenual cingulate cortex, a region implicated in depression and emotion regulation.
Taken together, these findings indicate that the neural representation of aesthetic musical emotions accords with an organisation into two basic dimensions of valence and arousal, similar to emotions evoked by other stimuli (Russell, 2003) and subserved by brain networks usually implicated in reward and vigilance functions, respectively. This adds further support to the notion that these emotions emerge from brain activity patterns partly shared with other emotions, including the more “basic” categories. However, these data also reveal a finer differentiation that is not accounted by traditional bi-dimensional models. More specific types of musical emotions appear to imply additional recruitment of other brain networks, outside those typically associated with emotion processing and the bidimensional construct of valence and arousal; in particular, these emotions also engage networks involved in motor, memory, self-reflective, and even visuo-spatial functions.
These data highlight the fact that emotion differentiation is partly determined by the cognitive processes concomitantly engaged with appraisals of value and motivational significance (Leventhal & Scherer, 1987). In addition, recent studies suggest that differences in emotional responses to music may also reflect changes in the functional connectivity (i.e. synchronization) of emotion-related areas such as the amygdala (Koelsch & Skouras, 2014) and striatum (Salimpoor, et al., 2013), with other distant brain regions including hippocampus, sensorimotor areas, medial prefrontal areas, or auditory cortex. This in turn also underscores the dynamic nature of emotional responses across distributed brain networks (Eryilmaz, Van De Ville, Schwartz, & Vuilleumier, 2011), rather than in terms of simple dimensions mediated by emotion-specific brain structures.
In sum, these findings converge to depict musical emotions as an embedding of multiple processes mediating sensory, motor, and cognitive functions (e.g. memory, attention) into a coordinated configuration of brain activation imbuing them with particular affective or motivational values. This in turn illustrates the fact that studies of the “complex” emotions induced by music may allow going beyond traditional models based on basic categories (Turner and Ortony, 1992) or polar dimensions (Russell, 2003). Moreover, this recruitment of sensorimotor and cognitive processes by music and concomitant emotions echoes the increasing amount of studies suggesting that music may yield significant benefits in training various cognitive and sensorimotor abilities, usefully exploited for rehabilitation in neurological patients or aging (Forsblom, Laitinen, Sarkamo, & Tervaniemi, 2009; Moore, 2013; Wan & Schlaug, 2010). A better understanding of the neural substrates of music-emotions may thus not only illuminate central questions about the nature of emotions in the brain but also open the way to novel and more efficient therapeutic applications.
Acknowledgments
The author’s work is supported by the Geneva Academic Society (Foremane Fund) and the Swiss National Science Foundation through grants to the Swiss Center for Affective Sciences (CISA) hosted by the University of Geneva. Special thanks to Wiebke Trost, Marcel Zentner, Carolina Labbé, and Natalia Fernandez for their contribution to projects described here.
References
Armony, J. L., & Vuilleumier, P. (2013). Handbook of human affective neuroscience. New York: Cambridge University Press.
Aube, W., Angulo-Perkins, A., Peretz, I., Concha, L., & Armony, J. L. (2014). Fear across the senses: brain responses to music, vocalizations and facial expressions. Soc Cogn Affect Neurosci.
Blood, A. J., & Zatorre, R. J. (2001). Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion. Proc Natl Acad Sci U S A, 98, 11818-11823.
Blood, A. J., Zatorre, R. J., Bermudez, P., & Evans, A. C. (1999). Emotional responses to pleasant and unpleasant music correlate with activity in paralimbic brain regions. Nat Neurosci, 2, 382-387.
Eryilmaz, H., Van De Ville, D., Schwartz, S., & Vuilleumier, P. (2011). Impact of transient emotions on functional connectivity during subsequent resting state: a wavelet correlation approach. Neuroimage, 54, 2481-2491.
Forsblom, A., Laitinen, S., Sarkamo, T., & Tervaniemi, M. (2009). Therapeutic role of music listening in stroke rehabilitation. Ann N Y Acad Sci, 1169, 426-430.
Gosselin, N., Peretz, I., Hasboun, D., Baulac, M., & Samson, S. (2011). Impaired recognition of musical emotions and facial expressions following anteromedial temporal lobe excision. Cortex, 47, 1116-1125.
Koelsch, S. (2014). Brain correlates of music-evoked emotions. Nat Rev Neurosci, 15, 170-180.
Koelsch, S., Fritz, T., DY, V. C., Muller, K., & Friederici, A. D. (2006). Investigating emotion with music: an fMRI study. Hum Brain Mapp, 27, 239-250.
Koelsch, S., Jentschke, S., Sammler, D., & Mietchen, D. (2007). Untangling syntactic and sensory processing: an ERP study of music perception. Psychophysiology, 44, 476-490.
Koelsch, S., & Skouras, S. (2014). Functional centrality of amygdala, striatum and hypothalamus in a “small-world” network underlying joy: an fMRI study with music. Hum Brain Mapp, 35, 3485-3498.
Leventhal, H., & Scherer, K. (1987). The relationship of emotion to cognition: A functional approach to a semantic controversy. Cognition and emotion, 1, 3-28.
Menon, V., & Levitin, D. J. (2005). The rewards of music listening: response and physiological connectivity of the mesolimbic system. Neuroimage, 28, 175-184.
Mitterschiffthaler, M. T., Fu, C. H., Dalton, J. A., Andrew, C. M., & Williams, S. C. (2007). A functional MRI study of happy and sad affective states induced by classical music. Hum Brain Mapp, 28, 1150-1162.
Moore, K. S. (2013). A systematic review on the neural effects of music on emotion regulation: implications for music therapy practice. J Music Ther, 50, 198-242.
Poldrack, R. A. (2006). Can cognitive processes be inferred from neuroimaging data? Trends Cogn Sci, 10, 59-63.
Russell, J. A. (2003). Core affect and the psychological construction of emotion. Psychol Rev, 110, 145-172.
Salimpoor, V. N., Benovoy, M., Larcher, K., Dagher, A., & Zatorre, R. J. (2011). Anatomically distinct dopamine release during anticipation and experience of peak emotion to music. Nat Neurosci, 14, 257-262.
Salimpoor, V. N., van den Bosch, I., Kovacevic, N., McIntosh, A. R., Dagher, A., & Zatorre, R. J. (2013). Interactions between the nucleus accumbens and auditory cortices predict music reward value. Science, 340, 216-219.
Trost, W., Ethofer, T., Zentner, M., & Vuilleumier, P. (2012). Mapping aesthetic musical emotions in the brain. Cereb Cortex, 22, 2769-2783.
Turner, T. J., & Ortony, A. (1992). Basic emotions: can conflicting criteria converge? Psychol Rev 99, 566-571.
Wan, C. Y., & Schlaug, G. (2010). Music making as a tool for promoting brain plasticity across the life span. Neuroscientist, 16, 566-577.
Zentner, M., Grandjean, D., & Scherer, K. R. (2008). Emotions evoked by the sound of music: characterization, classification, and measurement. Emotion, 8, 494-521.


