
 Stephan Hamann, Department of Psychology, Emory University
Stephan Hamann, Department of Psychology, Emory University
Recent decades have witnessed a knowledge explosion about all aspects of brain function. Neuroscience studies of emotion have also multiplied, using a wide array of methods from the molecular to the systems level across multiple species. Relatively recently, functional neuroimaging, primarily in the form of functional MRI (fMRI) has assumed a leading role in examining the brain basis of human emotion, with hundreds of papers published to date investigating a wide range of emotion phenomena. Substantial advances have been made in understanding the neural mechanisms involved in specific emotion domains, ranging from facial emotion processing to emotional memory. However, so far there has been surprisingly little high-level integration of affective findings across domains, and a coherent and organized consensus framework for understanding the neural underpinnings of emotion from the findings of neuroimaging studies has remained elusive.
In light of this, what have we actually learned from this proliferation of neuroimaging studies that illuminates fundamental aspects of emotion and their neural representation? What has neuroimaging added that would not have been known otherwise? Here I will focus on a few major ways that neuroimaging has contributed to the overall endeavor of understanding the emotional brain and highlight some challenges and future directions.
Many neuroimaging studies, including those of emotion, have focused on “brain mapping”, the mapping or association of brain functions to brain structures with the goal of elucidating the brain’s functional organization. But unlike a world map which has well-defined and universally recognized components and boundaries such as mountains and oceans, there is no similar agreement on the components and boundaries of maps of the emotional brain. Neuroimaging studies based on different psychological emotion views such as discrete basic emotions (e.g., fear), affective dimensions (e.g., arousal), or survival relevant circuits (e.g., defense) map essentially different theoretical constructs onto the brain, creating multiple model-dependent maps that complicate attempts to summarize across studies (see Figure 1 for discussion).
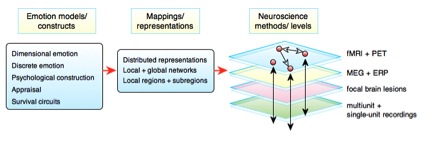
Neuroscience studies of emotion attempt to relate theoretical constructs drawn from emotion theories (fear, arousal, defense, etc.) to brain correlates at levels from brain systems to single neurons. These mappings of emotion constructs can take many forms, ranging from mappings at the level of brain regions or subregions (red spheres) to multi-region networks (connecting arrows). The functions of each region or network can be investigated via imaging and recording or lesion methods at multiple spatial scales (black arrows span analysis levels) and multiple time scales. Such mappings are complicated, however, by a lack of consensus regarding emotion models and by sometimes divergent findings obtained with different methods. For example, studies using direct electrical stimulation of the human amygdala, fMRI, and amygdala lesions each have reached substantially different conclusions about the emotional functions of the amygdala (Hamann, 2011). A major outstanding challenge is reconciling and integrating system-level theoretical models derived from functional neuroimaging results with other neuroscience methods at multiple levels
The process of mapping emotion in the brain is ultimately only as successful as these emotion models and their constructs, highlighting how theories of emotion play a critical role in determining the basic components and boundaries of maps charting the brain basis of emotions.
Despite these challenges, techniques such as neuroimaging meta-analysis have elucidated some key principles and have played an important role in recent theoretical debates (Hamann, 2012). For example, one major debate has focused on whether the separate emotions such as fear and disgust that are posited by discrete basic emotions theories actually map onto activation in unique brain regions in neuroimaging studies, as would be expected if each emotion had a dedicated neural circuit. If basic emotions are not reflected in brain activations, this would call into question the neural validity of such theories (Lindquist, Wager, Kober, Bliss-Moreau, & Barrett, 2012).
Neuroimaging meta-analyses can analyze associations between brain activation and emotion function across multiple studies. On the one hand, these analyses have found that basic emotions are indeed reflected in consistently greater activation in particular brain regions, allowing one to predict the brain regions likely to be activated during a particular emotion (Vytal & Hamann, 2010). However, meta-analyses have generally failed to find one-to-one mappings between emotions and brain regions, suggesting that it is generally invalid to infer a single emotion, basic or otherwise, from activation of an isolated region, despite the popularity of this type of inference (Poldrack, 2011; Stevens & Hamann, 2012). Instead, each brain region plays many different functional roles across multiple emotional situations, depending on the affective context and the broader network of other interacting regions active at the same time (Anderson, Kinnison, & Pessoa, 2013; Barrett & Satpute, 2013). A region such as the amygdala is like a highly versatile actor, best known for certain genres but able to play a wide range of roles depending on the ensemble of other actors interacting in a given scenario (e.g., fear, reward, arousal, novelty detection), rather than being like a typecast actor who plays the same role in every film (e.g., only fear).
These findings are not consistent with traditional basic emotion views that require dedicated emotion-specific brain regions that are not used for other emotions or other cognitive functions. However, they are consistent with theoretical variations on the basic emotions theme that allow for more flexible types of mappings between emotions and the brain, such as those that propose networks of brain regions as a more appropriate level of mapping between emotions and brain (Hamann, 2012). Preliminary evidence of the potential of network level analyses comes from fMRI studies that have used multivariate pattern classification methods, which can detect subtle patterns of activity reliably associated with mental states, even when those patterns are widely distributed across the brain. Studies using such methods have successfully distinguished and decoded multiple basic emotions from distributed patterns of brain activity, both within and across individuals, often in cases where standard fMRI analyses focused on individual regions fail to detect differences between emotions (Kassam, Markey, Cherkassky, Loewenstein, & Just, 2013; Peelen, Atkinson, & Vuilleumier, 2010).
Another major contribution of neuroimaging is its role in promoting new theoretical advances in affective science. fMRI occupies a special spatiotemporal niche among neuroscience methods, which enables the simultaneous investigation of activity across the entire brain, at multiple spatial scales, and at time scales well-suited for studying emotion. Powerful neuroimaging tools have also been developed for synthesizing and analyzing results from multiple studies in search for emergent patterns across studies. fMRI’s ability to record and analyze brain activation associated with emotion representations at multiple spatial and temporal scales facilitates the formulation and testing of alternative views of brain mechanisms of high-level emotion models. Although other methods such as neuropsychological lesion studies have also been fertile ground for theoretical development (Damasio, 2005), many recent theoretical advances and debates regarding emotion have centered on findings from neuroimaging, highlighting this method’s importance in driving theoretical change.
A promising theoretical development is the recent focus on identifying key adaptive survival challenges shared across species and the associated survival circuits that mediate a coordinated set of adaptive brain and behavioral responses (LeDoux, 2012). Survival circuits differ from basic emotions in that they are defined by brain circuits and adaptive functions that are conserved across mammals, rather than by subjective emotional experience (LeDoux, 2012). Although individual survival circuits do not map directly onto basic emotion categories, there are conceptual similarities between proposed circuits such as defense against harm and basic emotions such as anger and fear. Survival circuits have yet to be systematically investigated in human fMRI studies. A key question for such studies is whether survival circuits can be mapped consistently onto specific, evolutionarily conserved brain regions and networks.
Neuroimaging is ultimately only one method among many in the affective neuroscience toolbox, and like any method it has important limitations, such as limits on spatial and temporal resolution and its essentially correlational nature. The concept of converging operations (Bechtel, 2002) refers to the use of complementary evidence from multiple techniques or levels to corroborate experimental conclusions, overcoming weaknesses of individual techniques. A future challenge for affective science will be to apply this approach more systematically, to bridge and integrate across levels of analysis, brain organization, multiple methods and species. Initial efforts at such integration have shown that different methods can yield contradictory findings, highlighting the magnitude of the challenge ahead. Studies using permanent and reversible brain lesions to determine whether particular regions are critical for emotional function, versus merely correlated with it, will be particularly important in complementing neuroimaging’s correlational findings. Finally, another major challenge will be to achieve greater theoretical consensus regarding the representation and organization of emotion, which will promote synthesis across all levels of affective neuroscience and help to integrate multiple competing maps of the emotional brain into a common framework.
References
Anderson, M. L., Kinnison, J., & Pessoa, L. (2013). Describing functional diversity of brain regions and brain networks. NeuroImage, 73, 50–58. doi:10.1016/j.neuroimage.2013.01.071
Barrett, L. F., & Satpute, A. B. (2013). Large-scale brain networks in affective and social neuroscience: towards an integrative functional architecture of the brain. Current Opinion in Neurobiology, 23(3), 361–372. doi:10.1016/j.conb.2012.12.012
Bechtel, W. (2002). Aligning multiple research techniques in cognitive neuroscience: Why is it important? Philosophy of Science, 69(S3), S48–S58. doi:10.1086/341767
Damasio, A. (2005). Descartes’ Error: Emotion, Reason, and the Human Brain. Penguin.
Hamann, S. (2011). Introduction to the Special Issue on the human amygdala and emotional function. Neuropsychologia, 49(4), 585-588.
Kassam, K. S., Markey, A. R., Cherkassky, V. L., Loewenstein, G., & Just, M. A. (2013). Identifying Emotions on the Basis of Neural Activation. PLoS ONE, 8(6), e66032. doi:10.1371/journal.pone.0066032
LeDoux, J. (2012). Rethinking the Emotional Brain. Neuron, 73(4), 653–676. doi:10.1016/j.neuron.2012.02.004
Lindquist, K. A., Wager, T. D., Kober, H., Bliss-Moreau, E., & Barrett, L. F. (2012). The brain basis of emotion: A meta-analytic review. Behavioral and Brain Sciences, 35(03), 121–143.
Peelen, M. V., Atkinson, A. P., & Vuilleumier, P. (2010). Supramodal Representations of Perceived Emotions in the Human Brain. Journal of Neuroscience, 30(30), 10127–10134. doi:10.1523/JNEUROSCI.2161-10.2010
Poldrack, R. A. (2011). Inferring Mental States from Neuroimaging Data: From Reverse Inference to Large-Scale Decoding. Neuron, 72(5), 692–697. doi:10.1016/j.neuron.2011.11.001
Stevens, J. S., & Hamann, S. (2012). Sex differences in brain activation to emotional stimuli: A meta-analysis of neuroimaging studies. Neuropsychologia, 50(7), 1578–1593.
Vytal, K., & Hamann, S. (2010). Neuroimaging support for discrete neural correlates of basic emotions: a voxel-based meta-analysis. Journal of Cognitive Neuroscience, 22(12), 2864–2885.


Synapse transmission between neurons in different systems of dimension – how can it improve
the music imagination for beginners ?
Stepanov Ukraine
Considering the benefits of Math in a Music Educational Program
Kids of new generation should have different methods of training .They capable to perceive information faster, with cross-modal processing ,activating all senses at once : visual perception, audio analyzers , neuromotor functions.
http://educationinjapan.wordpress.com/2011/02/04/considering-the-benefits-of-digital-music-grammar-in-a-music-educational-program/